LC–MS as a Platform Method for mRNA CQA Analysis: Evaluating 5’ Capping Efficiency
Messenger RNA (mRNA) vaccines may continue to prove useful in combating new viral variants and more effectively controlling existing strains. However, the analytical approaches for quality control testing must keep pace to ensure the identity, safety, and efficacy of these new prophylactic and therapeutic modalities. This article describes a new approach for measuring critical quality attributes including the confirmation of 5’ cap structure and 5’ capping efficiency, as described herein.
The promise of messenger RNA (mRNA)‑based vaccines and therapeutics is being borne out today with the rapid development and emergency authorization of two mRNA vaccine therapeutics designed to fight the ongoing COVID-19 pandemic. Tozinameran (BNT162b2) from Pfizer‑BioNTech and mRNA-1273 from Moderna have demonstrated high efficacy against COVID-19 (1), and alternative vaccines are at various stages of development. The success of these two mRNA vaccines has led to a surge of interest, investments, and research into additional mRNA and nucleic acid‑based therapeutics. In many cases, the targets for these therapeutics are diseases that conventional drugs cannot treat. One such mRNA candidate currently in clinical trials is MRT5005 from Translate Bio, which is looking to treat cystic fibrosis (2).
As with all new modalities and as research into mRNA vaccines continues, improvements in areas such as drug delivery and design must occur during the creation of these vaccines (3). Unlike prior vaccines, which include a weakened or inactivated form of the pathogen to illicit immune response, mRNA vaccines provide a set of instructions that receiving cells then translate into a protein. In the case of the COVID-19 vaccines, the protein being produced is the “spike protein” that juts out from the surface of the corona virus. These spike proteins are subsequently recognized by our immune system as being foreign, triggering an immune response and the production of antibodies to neutralize the threat, the result being protection against future infection.
Throughout the process of creating an mRNA vaccine, regulatory requirements for the characterization of mRNA must be met. Though specific requirements from both the International Conference on Harmonization (ICH) and the U.S. Food and Drug Administration (FDA) have not yet been implemented, the FDA and the World Health Organization (WHO) have published guidelines for COVID‑19‑related vaccines and RNA-based diseases, respectively (4). The WHO draft discusses the evaluation of vaccines and calls out specific critical quality attributes (CQAs), particularly in identifying and quantifying proper cap structure and the poly(A) tail (5,6). Eukaryotic mRNA all contain a cap structure consisting of a 7-methyl guanosine connected to the first nucleotide of mRNA through a 5’ to 5’ triphosphate bridge. This capping is essential to many functions—for example, for recognizing the mRNA, promoting translation, adding resistance to exonuclease degradation, and more (7).
Depending on the method used to cap synthetic mRNA, different cap structure variants and precursors may be present. Methods to cap mRNA include those that rely on co‑transcriptional, post‑transcriptional, or enzymatic pathways. As a ubiquitous and highly efficient method, enzymatic preparation of capped mRNA is performed through in vitro transcription using a DNA template, which then produces mRNA with an uncapped triphosphate at its 5’ end. Typically,
the proper cap structure—Cap‑1 for higher eukaryotic species such as humans—can be enzymatically produced from the triphosphate species. Figure 1 details the process of producing Cap-1 from the triphosphate‑modified mRNA.


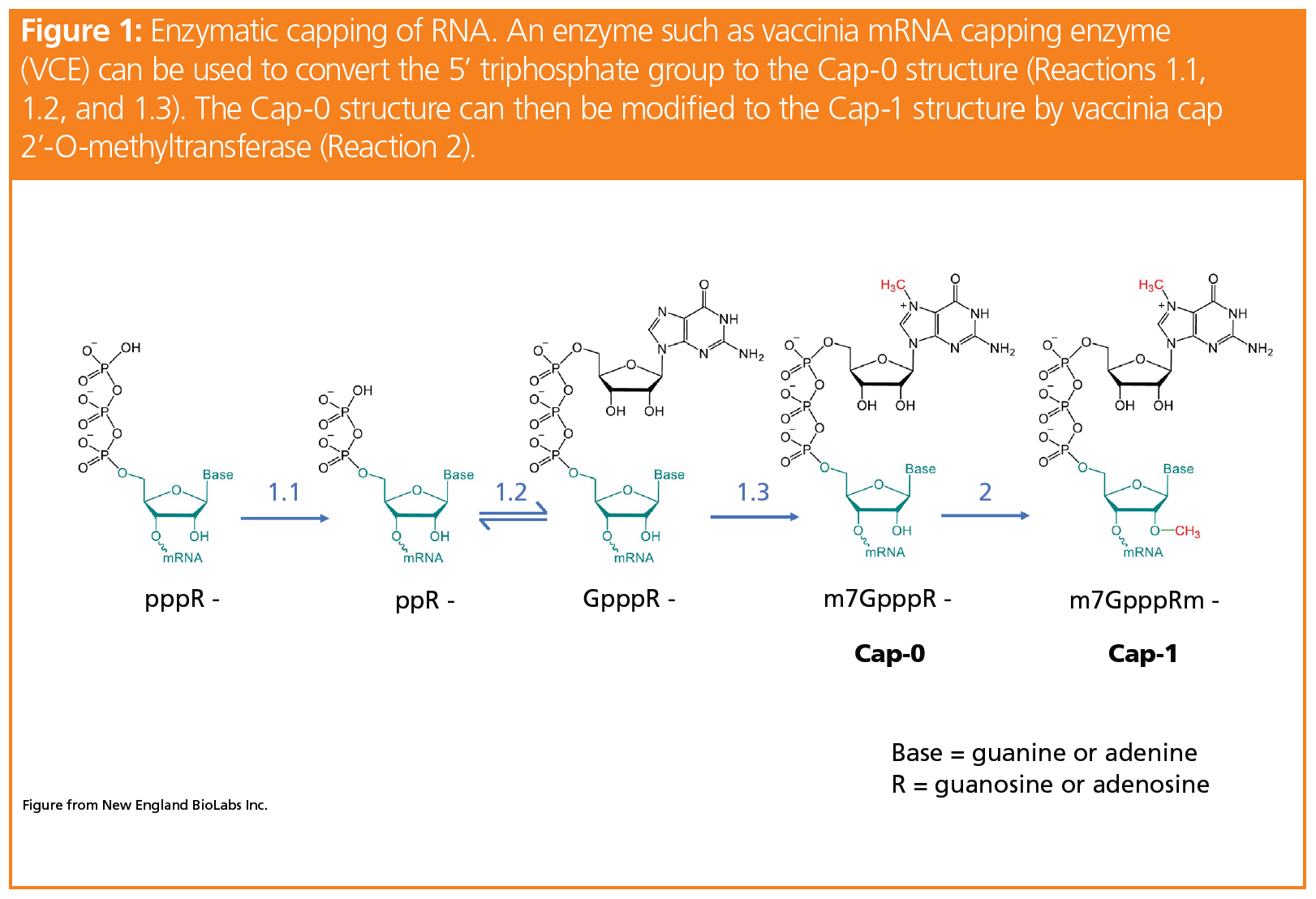
Though enzymatic capping typically produces high yields of Cap-1-modified mRNA, it can be seen from Figure 1 that there are four precursor cap structures throughout the process to form Cap‑1. Based on regulatory guidelines, the capping efficiency of the Cap-1 structure must be quantified and any impurities, such as the precursor cap structures, must be identified. Established techniques to analyze these CQAs, in particular the capping efficiency, include variations of enzymatic digestion coupled to gel electrophoresis or chromatography (8). In these methods, the digestion typically produces short strands of uncapped RNA or even mononucleotides to allow an estimation of the percentage of capped sample. However, these methods require the mRNA to be labelled, which adds another variable and lengthens the process.
To address this gap in analytics for unmodified mRNA, recent literature has demonstrated the benefits of liquid chromatography–mass spectrometry (LC–MS) analysis to this application, where the identification of non-Cap-1 species can be differentiated through differences in mass and retention time and quantified through MS after digestion of mRNA. Beverly and co-workers from the Novartis Institutes of Biomedical Research employed a biotinylated probe complementary to the 5’ end of mRNA RNAse H to catalyze the cleavage of pre-determined fragments from the 5’ end containing the cap structure. These fragments were enriched and analyzed via LC–MS. In addition, a more current publication in 2021 by Muthmann and co-authors performed digestion of the entire transcript as initiated by nuclease P1, snake venom phosphodiesterase, and dephosphorylation (9). With their method, they generated single nucleosides with or without the cap and quantified these products with triple quadrupole mass spectrometry.
Insights from these works and others show the benefits of implementing an LC–MS method versus an LC optical-only method when measuring CQAs of mRNA, including quantification of the 5’ cap fragments. For instance, quantification via UV relies heavily on the separation of the capped species and precursors, whereas LC–MS methods can unequivocally identify and quantify each component, even if not well separated, providing mass certainty as well. To this end, readily deployable, easy-to-use, and compliance‑ready LC–MS workflows may prove to be beneficial as the mRNA therapeutic pipeline surges.
Moreover, recently introduced LC technologies have been shown to improve the quantification of low-level impurities, for example the 5’ cap precursors leading up to Cap-1. This hybrid surface technology (HST) for column and LC systems hardware addresses and prevents nonspecific adsorption of analytes, particularly acidic compounds, to the electron-deficient metal surfaces of the LC fluid path (10). A highly cross-linked, ethylene-bridged siloxane polymer acts as a barrier that prevents analyte interactions with the metal surface. In particular, the surface technology has been shown to greatly improve the recovery of oligonucleotides, especially at lower level concentrations (10).
Example of 5’ Cap Structure Analysis with LC–MS
To demonstrate the benefits of LC–MS in combination with the new LC technologies available, we evaluated an unused hybrid surface column against an unused conventional stainless steel column for the analysis of synthetic RNA oligonucleotides using an ultrahigh‑pressure LC (UHPLC) system combined with a simple-to-use time‑of‑flight mass spectrometer (TOF‑MS). These synthetic oligonucleotides consist of 25 nucleotides of the same sequence but are modified with different groups on the 5’ end (pppR‑, ppR-, GpppR-, m7GpppR- or Cap-0, and m7GpppRm- or Cap-1). Figure 2 shows the separation and analyte peak of the Cap‑1 product using a mobile phase consisting of N, N – diisopropylethylamine (DIPEA) and 1,1,1,3,3,3-hexafluoroisopropanol (HFIP), which has been previously utilized for bioanalytical quantification of oligonucleotides due to its high resolution and MS signal intensities (11,12).

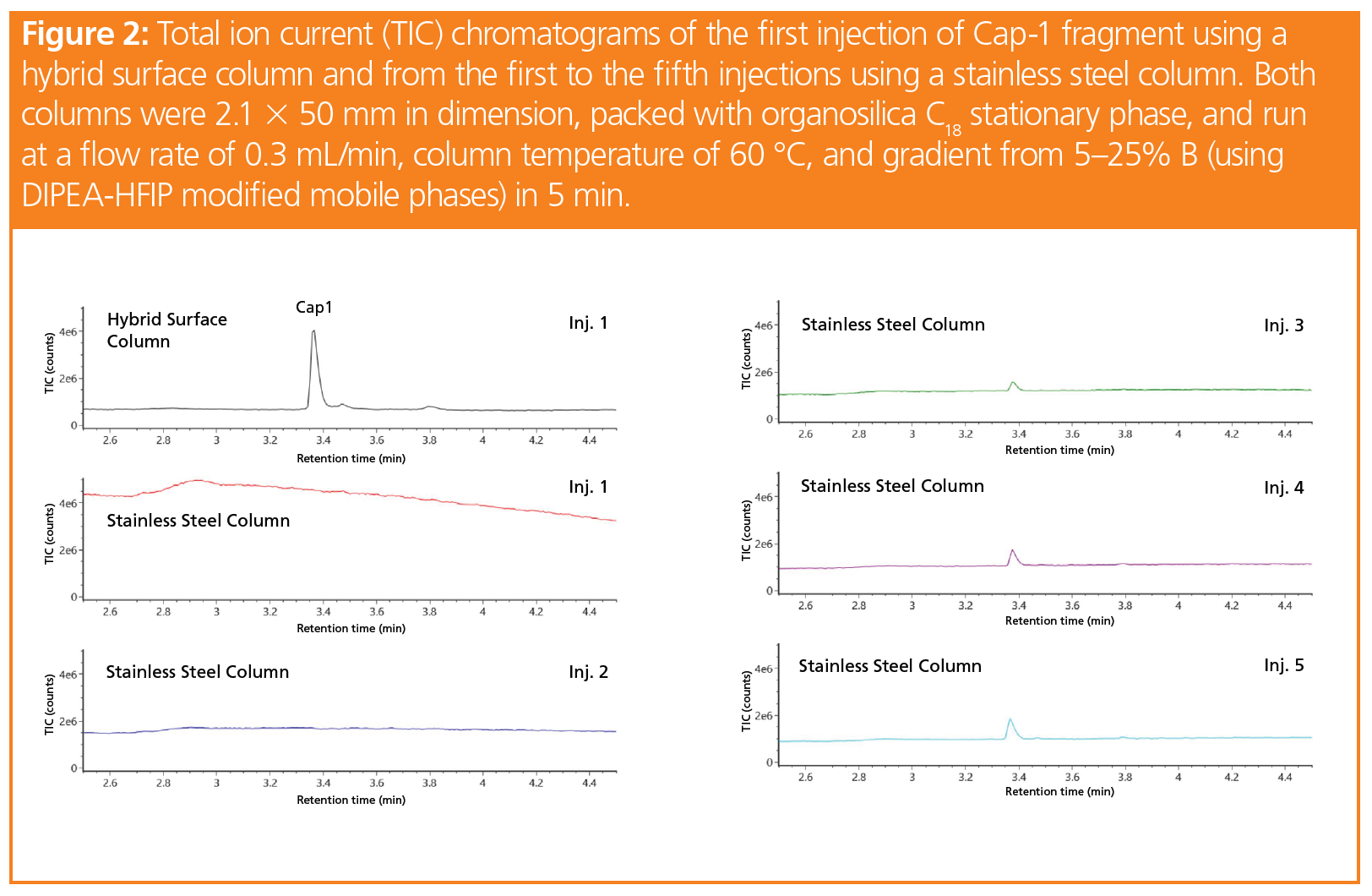
While only the first injection using the hybrid surface column is shown in Figure 2, the peak areas across five injections using this column were reproducible from injection to injection. In contrast, no peak is seen upon first injection using the stainless steel column. As the active sites on the stainless steel surface are slowly passivated by the sample from injection to injection, the Cap-1 peak is finally seen at injection 3. After injection 5, the peak area is still only 37% that of the hybrid surface column. The data suggest that employing inert LC components, such as hybrid surfaces, can improve the sensitivity and reproducibility of 5’ capping identification and quantification assays, and likely many other enzymatic assays where various substrates, precursors, and intermediates need to be tracked and understood.
Notably, from Figure 2, it can be seen that a sub-5-min method was developed to evaluate the capping efficiency of the Cap‑1 product. For oligonucleotide separations, historical methods have used longer gradients times. The method developed for the rapid elution of the 5’ capped species shown in Figure 2 employs accurate mass detection to identify the compounds and quantify them through various means, including the use of extracted ion chromatograms. With regards to real world samples, as opposed to the synthetically produced samples shown here, the biotinylated RNAse H probe would also need to be separated from the capped species for suitable quantitation. While not shown here, this rapid method can also easily separate a representative RNAse H probe from Cap-1.
To demonstrate the capability of this fast LC–MS method for measuring the capping efficiency of mRNA, we spiked samples of a Cap-1 solution with its four synthetic mRNA precursors (pppR-, ppR‑, GpppR-, and m7GppR-, also referred to as Cap-0) at dilution ratios of 1:10, 1:100, and 1:1000. The Cap-1 concentration was kept at 12.5 pmol/µL. The mass spectra for each dilution is shown in Figure 3. Here, the most abundant charge state for each species including Cap-1 was the [M-10H]10- charge state, which was confirmed by running each individual synthetic RNA fragment and is labelled in Figure 3.

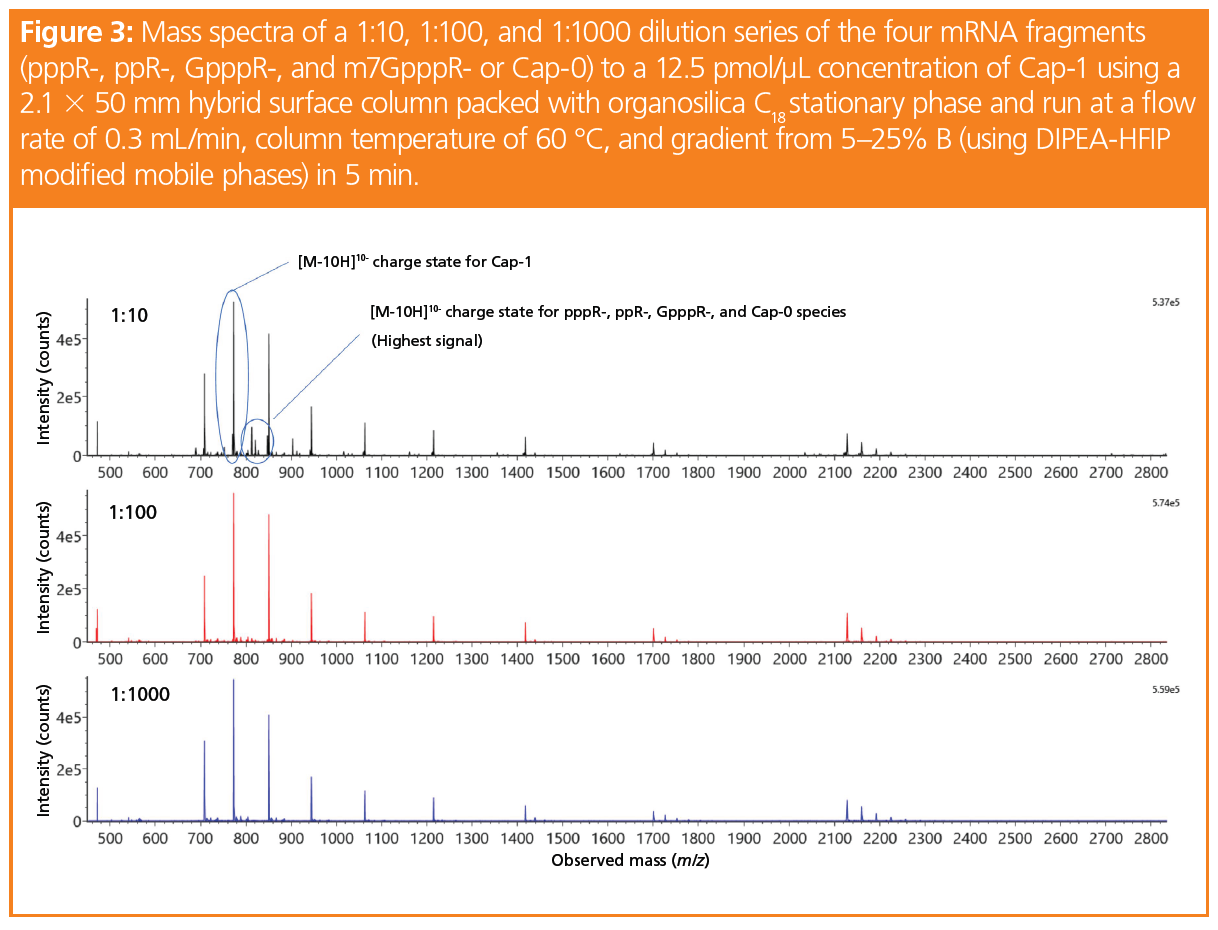
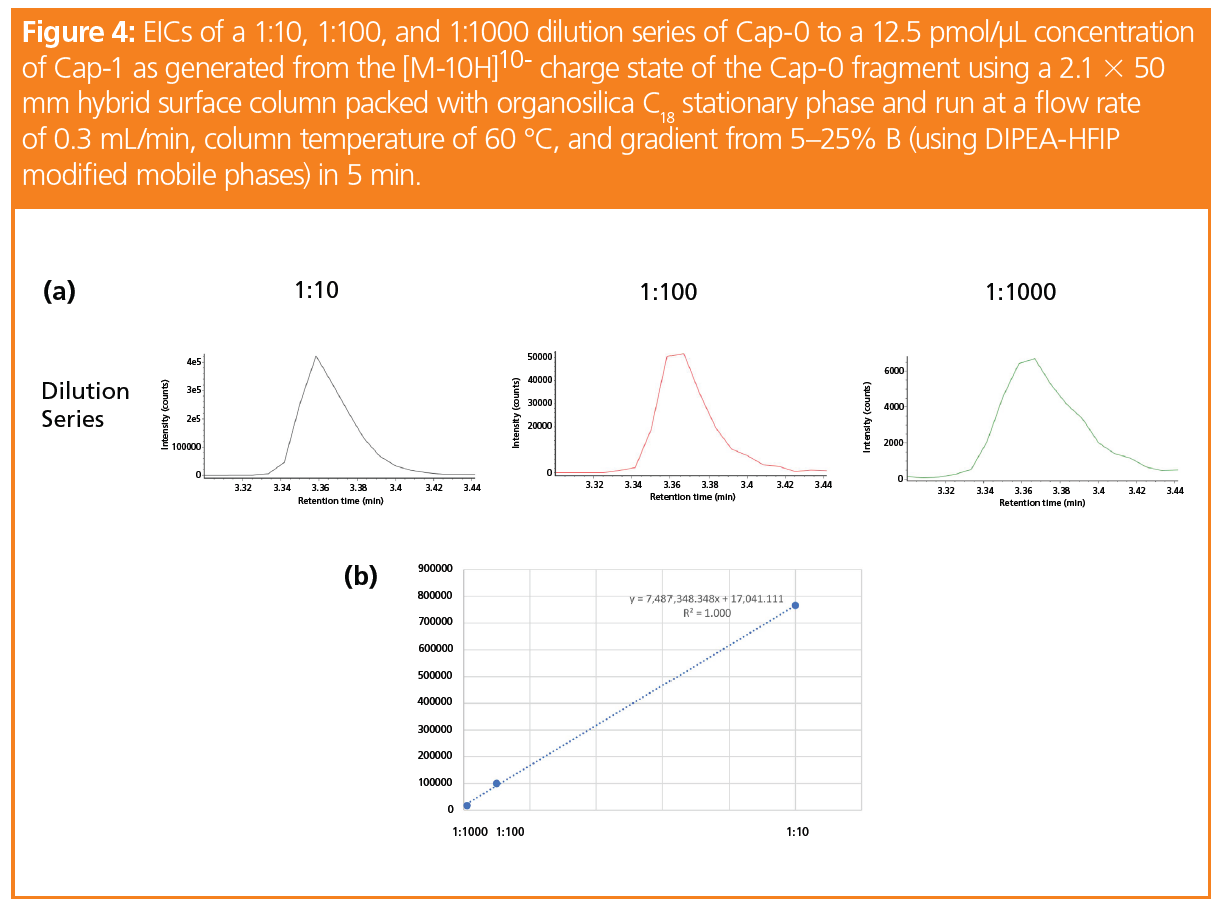
By taking the extracted ion chromatograms (EICs) of the entire isotope mass of each of the [M-10H]10- charge states for the five cap fragments (at 10 ppm), representative calibration curves for the four capped species used to generate Cap-1 can be produced. Even at mass loads of 12.5 fmol (as shown in the 1:1000 EIC for Cap-0 in Figure 4[a]), an EIC can be generated for integration. Figure 4(b) displays the calibration curve data for Cap-0 and its linear regression, with an R2 value of 1.000. Though not shown here, the linear regression for pppR‑, ppR-, and GpppR- also gave similar R2 values, which showcases the potential of MS quantitation for the evaluation of mRNA cap structures.

Conclusion
The rapid evaluation of the capping efficiency of mRNA described herein demonstrates the potential of LC–MS for mRNA analysis. Combined with an inert hybrid surface technology applied to the column, LC–MS was capable of assessing the 5’ capping of synthetic mRNA through the relative quantitation of a representative set of data. Coupled to an easy to use, compliance-ready LC–MS workflow, the 5-min method provides mRNA developers and manufacturers with a practical solution for the high-throughput analysis of 5’ cap structure. As the market for mRNA vaccines and gene therapy continues to expand, so will use of enzymatic assays to confirm critical quality attributes. The development and validation of these assays would benefit from compliance-ready LC–MS workflows that can be deployed across development, production, and quality control (QC) labs.
Acknowledgements
The work was supported by Martin Gilar, Weibin Chen, and Matthew A. Lauber at Waters Corporation as well as Siu-Hong Chan, Bijoyita Roy, and Brett Robb from New England BioLabs, Inc., who also provided the synthesized oligonucleotides.
References
- S.P. Teo, J. Pharm. Pract. 8971900211009650 (2021). doi:10.1177/08971900211009650
- M.Y.T. Chow, Y. Qiu, and J.K.W. Lam, Trends Pharmacol. Sci. 41, 715–729 (2020). doi:10.1016/j.tips.2020.08.002
- M. May, Nat. Med. 27, 930–932 (2021). doi:10.1038/s41591-021-01393-8
- D.J.A. Crommelin, T.J. Anchordoquy, D.B. Volkin, W. Jiskoot, and E. Mastrobattista, J. Pharma. Sci. 110, 997–1001 (2021). doi:10.1016/j.xphs.2020.12.006
- World Health Organization, Evaluation of the quality, safety and efficacy of RNA-based 5 prophylactic vaccines for infectious diseases: Regulatory Considerations (Geneva, Switzerland, 2020).
- I. Knezevic, M.A. Liu, K. Peden, T. Zhou, and H.N. Kang, Vaccines 9(2), 81 (2021). doi:10.3390/vaccines9020081
- A. Ramanathan, G.B. Robb, and S.H. Chan, Nucleic Acids Res. 44, 7511–7526 (2016). doi:10.1093/nar/gkw551
- I. Moya-Ramirez, C. Bouton, C. Kontoravdi, and K. Polizzi, Nucleic Acids Res. 48, e129 (2020). doi:10.1093/nar/gkaa955
- N. Muthmann, P. Spacek, D. Reichert, M. van Dulmen, and A. Rentmeister, Methods In Press (2021). doi:10.1016/j.ymeth.2021.05.018
- M. DeLano, T.H. Walter, M.A. Lauber, M. Gilar, M.C. Jung, J.M. Nguyen, et al., Anal. Chem. 93, 5773–5781 (2021). doi:10.1021/acs.analchem.0c05203
- T. Li, B. Huang, D. Li, Y. Zhu, L. Ding, and C. Shu, J. Chromatogr. B 1112, 61–66 (2019). doi:10.1016/j.jchromb.2019.02.027
- J. Liu, J. Li, C. Tran, K. Aluri, X. Zhang, V. Clausen, et al., Bioanalysis 11, 1967–1980 (2019). doi:10.4155/bio-2019-0137
Jennifer Nguyen is a Principal Scientist within Chemistry R&D at Waters Corporation. Since joining Waters in 2015, she has worked on developing new analytical technologies for both large and small molecules. Her current research centres on improving the characterization and quantitation of biopharmaceuticals and nucleic acid therapeutics. She has over 10 years of experience in LC–MS chromatographic separations of various molecules, with a focus on advancing new technologies.
E-mail: Jennifer_Nguyen@waters.com
Website: www.waters.com



















Regulatory Deadlines and Supply Chain Challenges Take Center Stage in Nitrosamine Discussion
April 10th 2025During an LCGC International peer exchange, Aloka Srinivasan, Mayank Bhanti, and Amber Burch discussed the regulatory deadlines and supply chain challenges that come with nitrosamine analysis.

