Hydrophobic Interaction Chromatography (HIC) for the Characterization of Therapeutic Monoclonal Antibodies and Related Products, Part 1: Theoretical Aspects


Hydrophobic interaction chromatography (HIC) is one of the historical analytical methods applied for the separation and purification of proteins. The main advantage of HIC is that separations are performed under mild—in most cases non-denaturing—conditions (ambient mobile phase temperature, aqueous mobile phase, and physiological pH). Thanks to the intense developments in the field of protein-based therapeutics, HIC has recently gained a huge interest and is currently considered as a reference method for the characterization of cysteine-linked antibody drug conjugates (ADCs) and widely used for the analysis of therapeutic monoclonal antibodies (mAbs) and bispecific antibodies (bsAbs), at both intact and sub-unit levels. In the first part of this two-part series, the current state of HIC is reviewed with a focus on retention and separation mechanism.
Hydrophobic interaction chromatography (HIC) is considered to be the reference technique to separate hydrophobic variants of mAbs and their subunits, and also to analyze the different populations of cysteine‐linked ADC molecules
that differ in their number of drugs per antibody, often referred to as the DAR (drug‐to‐antibody ratio) species (1,2,3). As well as the analytical characterization, HIC is also frequently used for protein purification during the downstream process (4). The main benefit compared to other LC modes (such as reversed‐phase liquid chromatography [RPLC], for example) is that HIC is assumed to be a non‐denaturing mode, therefore the native forms of the proteins are expected to be maintained. In addition, the separated species can be collected for further analysis, such as activity measurements. HIC is often employed as an orthogonal method to complement other chromatographic modes like ion‐exchange chromatography (IEX) or size‐exclusion chromatography (SEC) for protein characterization (5,6).
Despite the widespread use of HIC and reported publication of
many fundamental studies, the retention mechanism in HIC is not fully understood and none of the proposed theories has received general acceptance (3). Different interpretations are often mentioned in the literature, including hydrophobic interaction, hydrophobic effects, “long‐range non‐hydrophobic” interactions, solvophobic effects, salting‐out, dehydration of proteins or structural rearrangement like reversible and irreversible conformational changes of proteins. In the last few years, several fundamental studies helped to better understand HIC mechanism and thus to develop more robust methods. The aim of this two‐series article is to review the latest findings in HIC, to summarize the new developments in HIC column technology and present some recent applications from the field of therapeutic protein analysis including uni‐ and multi‐dimensional separations. In this first part the focus is on theoretical aspects.
Retention in HIC
To retain proteins in HIC, kosmotropic salts are typically added to the mobile phase. These salts generally reduce protein solubility, while stabilizing protein structure and facilitate reversible interactions between the protein’s nonpolar (more hydrophobic) surfaces and the stationary phase’s hydrophobic functional ligands. As a result, protein binding occurs at
high salt concentration, while elution takes place at low salt concentration (3,4,7,8,9). Then, the separation is based on reversible partitioning of proteins between a mobile phase containing the salt (such as ammonium sulfate) and a mildly hydrophobic stationary phase (for example, a butyl phase with low ligand density). Normally, increasing the salt concentration increases partitioning toward the stationary phase, while reducing the salt concentration increases partitioning toward the mobile phase. Because of the strong dependence of protein retention on salt concentration (“on‐off”, or “bind and elute” like mechanism), HIC is often operated in gradient mode, starting at a high salt concentration and ending with a low salt concentration (often called inverse salt gradient) (10). Figure 1 illustrates the strong impact of salt concentration (M) on the retention (k) of some model proteins (11).


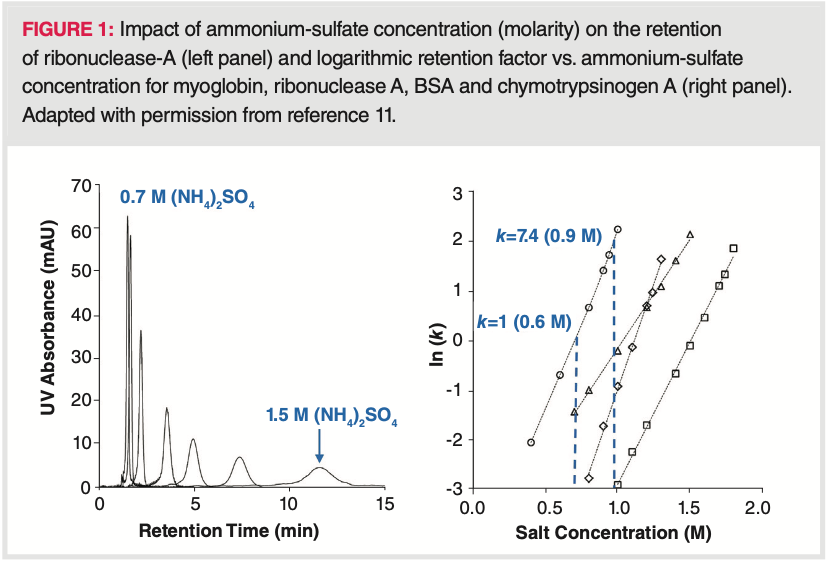
Chromatographic separation based on hydrophobic interactions was first reported by Tiselius (12). He used the term “salting‐out chromatography." The salting‐out effect is based on electrolyte–nonelectrolyte interactions (corresponding to mobile phase‐protein interaction in HIC), in which the nonelectrolyte becomes less soluble at high salt concentrations. The retention of proteins is mainly explained by the formation and collapsing of water layers surrounding the proteins and stabilized through H‐bonding. When adding salts into the mobile phase, water molecules solvate the salt ions predominantly. Therefore, the number of water molecules available to interact with the hydrophilic part of the protein will decrease. Under these conditions, the protein–protein and protein‐ligand intermolecular interactions become stronger due to the decreased amount of surrounding water molecules and proteins will be retained (3). Later on, several terms were used to describe such types of separations, including “hydrophobic chromatography”, “hydrophobic affinity chromatography (HAC)”, “hydrophobic interaction chromatography (HIC)”, “salt mediated separation of proteins” or “salt‐promoted adsorption chromatography (SPAC)”.
The solvophobic theory is also often used to explain HIC retention. It describes the interactions between a polar solvent (aqueous mobile phase) and less polar solute (protein). As a result of H‐bonding and other polar interactions, strong cohesive forces exist between the solvent molecules and provide a strongly structured order for the solvent (13). Therefore, less polar solutes tend to be insoluble due to the strong solvent–solvent binding interactions. According to this, the solute molecules bind to the surface of the stationary phase due to their rejection from the solvent and their affinity for the hydrophobic stationary phase. So, the retention is partly explained by the interactions between the solute and stationary phase and partly by the rejection of solute from the mobile phase solvent.
An entropy‐driven model can also describe the retention in HIC. The structure of water is highly ordered thanks to dipole–dipole interactions in a three‐dimensional structure, determined by the high surface tension of water. If hydrophobic moieties are dissolved in an aqueous system, the neighbouring water molecules have to be separated from each other thus forming a “cavity” for the less polar solute (14). This requires the investment of energy, depending on the surface area of the cavity and the surface tension. If two or more solutes (or solute and ligand) are associated, their hydrophobic contact surface area is reduced, and therefore energy is released. In other words, the interaction between hydrophobic molecules in aqueous solutions occurs spontaneously and is driven by the system’s entropy change (15,16,17). However, it has been shown that hydrophobic interactions are entropy driven at low temperatures, but enthalpy driven at elevated temperatures (18). Now it seems, that under practical conditions, both enthalpy and entropy control take place. It explains that in some conditions, retention increases with temperature while for other cases retention may decrease with temperature.
Is HIC Indeed Non-denaturing and Non-destructive?
HIC is often referred to as a non‐denaturing and non‐destructive chromatographic mode. Over the years, several studies have nevertheless proven that conformational changes may occur during the adsorption of proteins onto hydrophobic surfaces (19,20,21,22). Now, it seems that a certain fraction of the injected protein unfolds upon adsorption onto HIC stationary phases resulting in stronger binding and delayed elution, which can explain the loss of protein recovery observed in some cases (23,24,25). The partial unfolding of the proteins has been experimentally proven and a reversible unfolding mechanism was recently proposed (26). The rate of unfolding upon adsorption depends both on the type of stationary phase and on the protein properties. It was observed that “softer” proteins (which possess a more compressible structure) are more prone towards unfolding and showed higher retention (27). Based on melting point measurements (using differential scanning calorimetry), it could be shown that the conformational changes occurring in an antibody or antibody fragments upon adsorption onto HIC surface are directly proportional to the hydrophobicity of the stationary phase (28). Then upon elution (when protein is released from the HIC stationary phase), the protein takes back its original (native) conformation. It was also observed that antibody fragments which are less stable, are more likely to undergo conformational changes upon adsorption.
A model was recently developed, accounting for protein unfolding and intermolecular interactions in the adsorbed state (29). For the intermolecular interactions, we can distinguish so‐called preferred and multilayer adsorption on the adsorbent surface, as well as exclusion effects due to size exclusion and electrostatic repulsion. The mechanism of protein binding and spreading on a HIC surface is shown in Figure 2. First, the protein adsorbs on a free active site on the adsorbent surface in its native form and then, the protein‐binding surface spreads by attaching other active sites which results in unfolding. Additional protein molecules try to bind through positive cooperative adsorption, preferred adsorption, and multilayer adsorption. For proteins possessing unstable structures, the overall adsorption is dominated by the binding of unfolded species at low surface coverage and by positive cooperative adsorption at high surface coverage. Furthermore, exclusion effects strongly influence adsorption equilibrium, particularly at low surface coverages. So in the end, simultaneously, several different equilibrium processes (that is, adsorption, desorption, unfolding, folding, positive cooperative adsorption, and positive cooperative desorption) determine the retention in HIC.

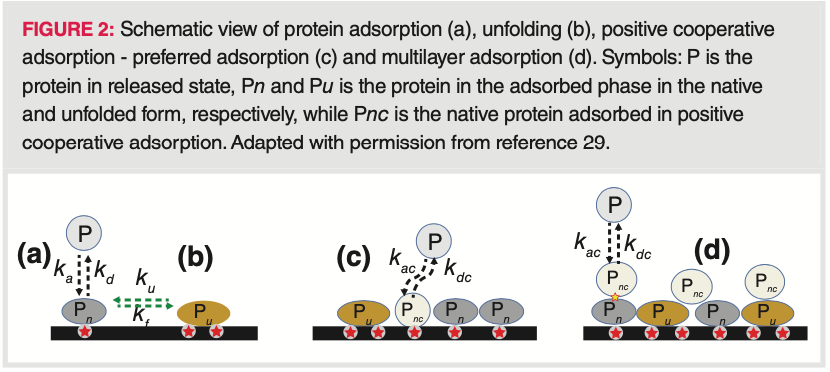
The strength of protein binding depends on the “footprint” of the protein, this is why aggregates are more retained than monomeric forms. Besides the size of the binding footprint, interactions between the protein surface and the stationary phase are likely to play an important role because they determine the preferred binding orientation. It has been demonstrated that even a small protein (for example, lysozyme) can interact with the chromatographic surface in orientation‐specific ways, depending on various factors such as injected amount/volume, mobile phase pH, and resin structure (30,31). It has also been shown recently that for antibodies, the various mAb domains interact differently with the stationary phase, each contributing to the overall binding strength and selectivity. In practice, it can be assumed that a distribution of different binding states and binding orientations exists for most proteins with the most favourable binding configuration being the most probable determining the overall retention (32).
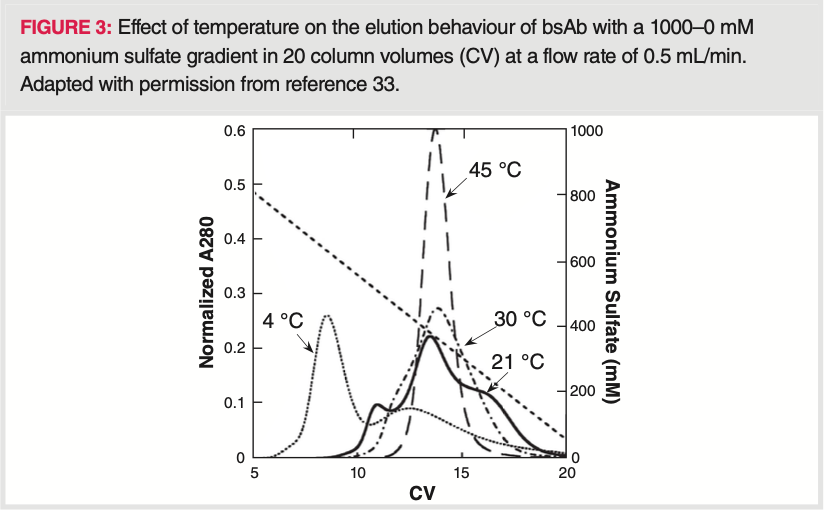
Since proteins can be oriented in different ways on the HIC stationary phase and can unfold upon binding in different extents (depending on their orientation, stationary phase hydrophobicity, temperature and pH), they can lose their secondary structure. The change in secondary structure can result in multiple peak elution. Some examples have been reported earlier, such as two‐peak elution of α‐lactalbumin, bovine serum albumin, and of β‐lactoglobulin on various HIC resins (20,22,23). Very recently, Carta et al. showed a multiple (three) peak elution of bivalent bispecific antibodies from HIC phases (33). While three peaks were obtained at short residence times (binding time) and room temperature, residence times longer than 27 min or operating at 45 °C resulted in a single merged peak, indicating that the underlying mechanism occurs on time scales comparable to that of a chromatographic separation. Holding the protein on the resins prior to elution enriched the late‐eluting peak, indicating that multiple binding states formed on the chromatographic surface are responsible for this behaviour. The multiple and merged peak elution of the bsAb is shown at various temperatures on Figure 3.

It has also been shown that the rate of conformational changes upon adsorption in HIC depends on sample load (injection, feed) too (26). The unfolding phenomenon seems
to enhance when increasing the salt content, and to decrease when increasing column loading. The latter phenomenon was explained by the reduction of available adsorption sites which aroused from steric hindrance to mass transport at the solid interface.
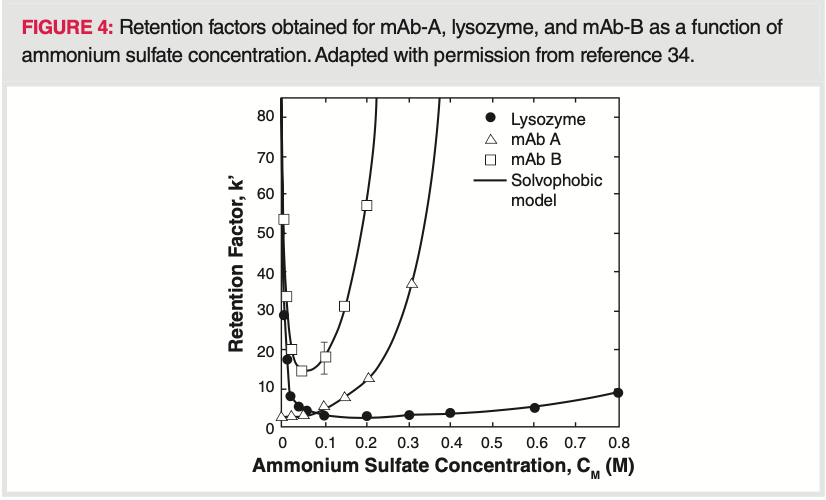
As discussed, protein retention is normally reduced by decreasing the salt concentration. However, in some cases, protein retention may increase at low salt concentrations resulting in a U‐shaped retention factor curve (retention versus. salt concentration) (34). It has been shown that some proteins may elute partially in gradient mode and partially trapped in the column. Solvophobic effects and other adsorptive interactions can lead to such U‐shaped retention factor curves in HIC. As an example, hydrophobic binding and protein unfolding on the stationary phase surface can lead to strong retention at low salt concentrations, weaker interactions at intermediate salt concentrations and strong binding again at high salt concentration. Moreover, U‐shaped retention factor curves can be the result of interplay between hydrophobic and electrostatic interactions (35,36,37). MAbs can also exhibit U‐shaped retention factor curves and a critical gradient slope can be determined beyond which 100% recovery is no longer possible (34). Figure 4 shows retention factor curves, obtained for two mAbs and lysozyme. Elution with a reverse gradient is also demonstrated at low salt concentrations for these proteins. Understanding this behaviour is important to design gradient HIC separations appropriately, since gradient slope may impact the recovery.

Conclusion
This article summarizes the most recent information that can be found in literature about hydrophobic interaction chromatography (HIC), which is widely applied for the analysis and purification of proteins. The first part of the two‐series articles focuses on theoretical aspects, particularly on retention mechanisms observed in HIC, with the goal to develop more robust methods. Various contributions to retention have been discussed including hydrophobic interaction, hydrophobic effects, “long‐range non‐hydrophobic” interactions, solvophobic effects, salting‐out, dehydration of proteins or structural rearrangement
like reversible and irreversible conformational changes of proteins.
HIC is often considered as a non‐denaturing chromatographic technique, which is advantageous for protein analysis. This article is also intended to decipher whether HIC can indeed be considered as a non‐denaturing and non‐destructive technique.
References
1) B. Wiggins, L.L. Shin, H. Yamaguchi, and G. Ratnaswamy, J. Pharm. Sci. 104, 1362–1372 (2015).
2) M. Haverick, S. Mengisen, M. Shameem, and A. Ambrogelly, mAbs 6, 852–858 (2014).
3) S. Fekete, J.L. Veuthey, A. Beck, D. Guillarme, J. Pharm. Biomed. Anal. 130, 3–18 (2016).
4) J.C. Janson, Protein purification: Principles, High Resolution Methods, and Applications (John Wiley & Sons, New Jersey, USA, 3rd ed., 2011).
5) J.A. Queiroz, C.T. Tomaz, and J.M.S. Cabral, J. Biotech. 87, 143–159 (2001).
6) A. Rodler, R. Ueberbacher, B. Beyer, and A. Jungbauer, Prep. Biochem. Biotechnol. 49, 1–20 (2019).
7) G. Carta and A. Jungbauer, Protein Chromatography: Process Development and Scale-Up (Wiley-vch Verlag, Weinheim, Germany, 2010).
8) P. Gagnon, Purification Tools for Monoclonal Antibodies (Validated Biosystems, Tucson, Arizona, USA, 1996).
9) F.E. Regnier, Science 222, 245–252 (1983).
10) W.R. Melander, D. Corradini, and C. Horvath, J. Chromatogr. 317, 67–85 (1984).
11) M. Baca, J. De Vos, G. Bruylants, K. Bartik, X. Liu, K. Cook, and S. Eeltink, J. Chromatogr. B. 1032, 182–188 (2016).
12) A. Tiselius, Mineral Geol. 26B, 1–5(1948).
13) C. Horváth, W. Melander, and I. Molnár, J. Chromatogr. 125, 129–156 (1976).
14) I. Molnár, Chromatographia 62, S7–S17 (2005).
15) S. Lewin, Displacement of Water and Its Control of Biochemical Reactions (Academic Press, New York, USA, 1974).
16) H.P. Jennissen, Biochemistry 15, 617–642 (1976).
17) J.L. Ochoa, Biochimie 60, 1–15 (1978).
18) R.L. Baldwin, Proc. Natl. Acad. Sci. U.S.A. 83, 8069–8072 (1986).
19) T.T. Jones and E.J. Fernandez, J. Colloid Interface Sci. 259, 27–35 (2003).
20) A. Jungbauer, C.Machold, and R. Hahn, J. Chromatogr. A. 1079, 221–228 (2005).
21) Y. Xiao, T.T. Jones, A.H. Laurent, J.P. O’Connell, T.M. Przybycien, and E.J. Fernandez, Biotechnol. Bioeng. 96, 80–93 (2007).
22) R.W. Deitcher, J.P. O’Connell, and E.J. Fernandez, J. Chromatogr. A. 1217, 5571–5583 (2010).
23) E. Haimer, A. Tscheliessnig, R. Hahn, and A. Jungbauer, J. Chromatogr. A. 1139, 84–94 (2007).
24) R. Ueberbacher, E.Haimer, R.Hahn, and A. Jungbauer, J. Chromatogr. A. 1198–1199, 154–163 (2008).
25) J.A. Queiroz, C.T. Tomaz, and J.M.S. Cabral, J. Biotechnol. 87, 143–159 (2001).
26) R. Muca, W. Marek, M. Zurawski, W. Piatkowski, and D. Antos, J. Chromatogr. A. 1492, 79–88 (2017).
27) B.C.S. To and A.M. Lenhoff, J. Chromatogr. A. 1141, 191–205 (2007).
28) B. Beyer and A. Jungbauer, J. Chromatogr. A. 1552, 60–66 (2018).
29) R. Muca, M. Kołodziej, W. Piatkowski, G. Carta, and D. Antos, J. Chromatogr. A. 1625, 461309 (2020).
30) F. Dismer, M. Petzold, and J. Hubbuch, J. Chromatogr. A. 1194, 11–21 (2008).
31) J. Kittelmann, K.M.H.Lang, M.Ottens, and J. Hubbuch, J. Chromatogr. A. 1482, 48–56 (2017).
32) J. Robinson, D. Roush, and S. Cramer, J. Chromatogr. A. 1563, 89–98 (2018).
33) L.K. Kimerer, T.M. Pabst, A.K. Hunter, and G. Carta, J. Chromatogr. A. 1617, 460836 (2020).
34) A. Creasy, J. Lomino, G. Barker, A. Khetan, and G. Carta, J. Chromatogr. A. 1547, 53–61 (2018).
35) W. Melander, Z. El Rassi, and C. Horvath, J. Chromatogr. 469, 3–27 (1989).
36) B.K. Nfor, M. Noverraz, S. Chilamkurthi, P.D.E.M. Verhaert, L.A.M. van der Wielen, and M. Ottens, J. Chromatogr. A. 1217, 6829–6850 (2010).
37) Y.F. Lee, H. Graalfs, and C.Frech, J. Chromatogr. A. 1464, 87–101 (2016).
AUTHORS
Szabolcs Fekete is a Scientific Collaborator at the University
of Geneva, Switzerland, in the Analytical Pharmaceutical Chemistry group. He was the awardee of the LCGC Emerging Leader Award in Chromatography in 2020.
Amarande Murisier is a PhD student at the University of Geneva, in Switzerland. Her PhD thesis focuses on novel chromatographic and electrophoretic techniques for the analysis of therapeutic proteins in the group of Jean-Luc Veuthey and Davy Guillarme.
Davy Guillarme is a Senior Lecturer and Research Associate
at the University of Geneva, in Switzerland. He is also an editorial board member of LCGC Europe.
Koen Sandra is the editor of “Biopharmaceutical Perspectives”. He is CEO at RIC group (Kortrijk, Belgium) and Visiting Professor at Ghent University (Ghent, Belgium). He is also a member of LCGC Europe’s editorial advisory board. Direct correspondence about this article to the Editor-in-Chief, Alasdair Matheson: amatheson@mjhlifesciences.com


 for the Characterization of Therapeutic Monoclonal Antibodies and Related Products, Part 1: Theoretical Aspects")












Common Challenges in Nitrosamine Analysis: An LCGC International Peer Exchange
April 15th 2025A recent roundtable discussion featuring Aloka Srinivasan of Raaha, Mayank Bhanti of the United States Pharmacopeia (USP), and Amber Burch of Purisys discussed the challenges surrounding nitrosamine analysis in pharmaceuticals.



